Polygenní dědičnost
(přesměrováno z Aditivní model polygenní dědičnosti)


Při polygenní dědičnosti se určitý znak dědí ve více genech, z nichž každý má na fenotyp jen malý vliv. Tyto geny o malém účinku označujeme jako minor geny. Znaky děděné tímto způsobem bývají většinou měřitelné (např. výška, váha, …), proto se používá synonymní název kvantitativní genetika.
Složitější případ, kdy dochází k interakcím a zároveň je fenotyp ovlivňován vnějším prostředím, označujeme termínem multifaktoriální dědičnost. Při tomto typu dědičnosti často polygenní systém interaguje i s geny většího účinku – major geny.
V praxi se pojmy „polygenní“ vs. „multifaktoriální“ dědičnost často (avšak ne zcela správně) používají bez zdůraznění jejich rozdílnosti. Multifaktoriální dědičnost je jednoduše princip, kde se kombinuje vliv složky zevního prostředí a vliv genetické složky – představované právě polygenním systémem (jak je vysvětleno dále).
Genetický princip[upravit | editovat zdroj]
Rozptyl fenotypu při polygenní dědičnosti má tři hlavní složky: složku genetickou (VG), složku vnějšího prostředí (VE) a interakce genotypu a prostředí (VGE).

Poměr míry rozptylu fenotypu závislé na genetických faktorech a faktorech vnějšího prostředí určuje heritabilita.

VG dále dělíme na složky rozptylu způsobené aditivní složkou (VA), působením dominance mezi alelami (VD) a nealelními interakcemi (VI).
Pro základní pochopení principů polygenní dědičnosti uvažujeme zjednodušený aditivní model, který pomůže pochopit složitější multifaktoriální interakce.
Aditivní model polygenní dědičnosti[upravit | editovat zdroj]
Tento zjednodušený model předpokládá v rámci genu alelickou interakci zvanou semidominance. Při té dochází k tomu, že je hodnota fenotypu heterozygota průměrem hodnot fenotypů homozygotů. Dále mezi jednotlivými geny platí nealelní interakce typu aditivita (kumulativnost). Jejím důsledkem je to, že se účinky jednotlivých genů sčítají. Ve výsledku tedy záleží pouze na počtu dominantních a recesivních alel nezávisle na poloze v genomu a vzájemné konfiguraci.
Jednotlivé alely označujeme dolním indexem podle příslušnosti ke genu. Vzhledem k tomu, že pro fenotyp je určující počet aktivních alel, jedinci s genotypem A1A1A2a2a3a3 mají stejný fenotyp jako jedinci s genotypem A1a1A2a2A3a3.
Fenotypové rozptyly při křížení[upravit | editovat zdroj]
V následujících odstavcích bude polygenní dědičnost vysvětlena na zjednodušeném příkladu výšky populace, kdy je výška děděna třemi geny a veškerý vliv prostředí je vyloučen. Genotyp AAAAAA určuje výšku 200 cm a genotyp aaaaaa výšku 150 cm. Heterozygot ve všech genech (AaAaAa) bude mít stejnou výšku jako genotyp AAAaaa z výše popsaných důvodů.
Čistě polygenní dědičnost je v přírodě výjimkou, častěji je fenotyp ovlivňován i vnějším prostředím. Proto i fenotypy nelze začlenit do jednotlivých skupin s přesnou velikostí, jako je předpokládáno v následujícím textu čistě jen podle genomu, ale jejich výška se rozloží mezi jednotlivé odhady výšky na základě genotypu. Grafem závislosti množství jedinců na výšce tedy nebudou pouze sloupce nad určitými výškami jako níže, ale křivka s vrcholy v daných místech.
Cross[upravit | editovat zdroj]
Při křížení dominantního a recesivního homozygota mají všichni jedinci F1 generace stejný genotyp – polovinu dominantních alel od jednoho rodiče a druhou polovinu recesivních alel od druhého rodiče. (Jsou ve všech genech heterozygotní – AaAaAa.) Proto bude jejich fenotyp projevem mezi rodičovskými fenotypy. Při křížení vysoké populace P1 a malé populace P2 bude mít každý jedinec F1 populace stejný genotyp a tedy velmi malý (resp. při úplném zanedbání vnějších vlivů žádný) rozptyl fenotypu, protože mají každý tři aktivní alely. Fenotypový rozptyl populace F1 lze proto použít pro výpočet heritability.
Intercross[upravit | editovat zdroj]
Při křížení F1 generace mohou potomci zdědit všechny možné kombinace alel ve všech třech genech, protože oba rodiče mají jak dominantní, tak recesivní alelu pro každý gen. Jejich genotypy budou mít tedy 0 až 6 aktivních alel (aaaaaa, Aaaaaa, …, AAAAAA). Proto se i rozptyl fenotypu bude pohybovat mezi fenotypy obou parentálních generací (výška 150–200 cm). Nejčastěji budou zastoupeny fenotypové projevy vprostřed a nejméně často oba extrémy, rozložení fenotypu bude tedy podle Gaussovy křivky.
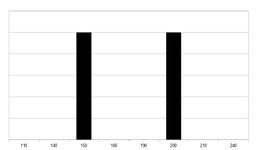
Parentální generace P – dvě odlišně vysoké homozygotní generace.
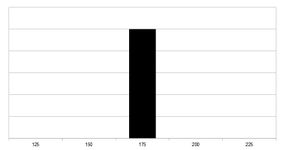
Filiální generace F1 – jedna čistě heterozygotní generace s uniformní výškou.
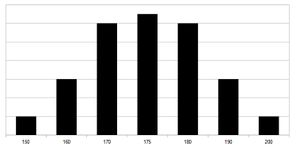
Filiální generace F2 – Fenotyp rozdělen do hypotetických 7 skupin podle genotypu (viz text). Rozložení odpovídá Gaussově křivce.



Polygenní dědičnost s prahovým efektem[upravit | editovat zdroj]

Náchylnost ke genetické vadě může být děděna polygenně tím způsobem, že určitý limitní počet recesivních alel způsobí vznik onemocnění. V tomto případě hovoříme o prahovém efektu polygenní dědičnosti. Je-li náchylnost k chorobě děděna ve třech genech, jedinec bude postižen například při 5 recesivních alelách (Aaaaaa) a více.
Odkazy[upravit | editovat zdroj]
Související články[upravit | editovat zdroj]
- Multifaktoriální dědičnost
- Alela
- Heritabilita
- Autozomálně dominantní a autozomálně recesivní dědičnost
- Nemendelovská dědičnost
Použitá literatura[upravit | editovat zdroj]
- OTOVÁ, Berta, et al. Lékařská biologie a genetika I. díl. 1. vydání. Praha : Karolinum, 2008. 123 s. s. 21–26. ISBN 978-80-246-1594-3.